Dott. Massimo Crimi - Facoltà di Scienze
- Università degli Studi di Verona
ATTIVITA’ DIDATTICA
Dal 1997 ad oggi:
Assistenza didattica per le
esercitazioni (laboratori sperimentali) del corso di
Biochimica-Biochimica Vegetale, corso di laurea in Biotecnologie
Agro-Industriali;
Componente ufficiale delle
commissioni di esame per i corsi di Biochimica-Biochimica Vegetale,
Fisiologia Vegetale, Biologia Molecolare Vegetale ed Ecofisiologia
Vegetale; corso di laurea in Biotecnologie Agro-Industriali;
Componente ufficiale della
commissione per la prova di conoscenza della lingua Inglese
Componente delle commissioni di
Laurea in Biotecnologie Agro-Industriali;
Rappresentante dei Ricercatori
della Facoltà di Scienze al Senato Accademico dell’Ateneo
di Verona
Componente della Commissione
Permanente per la Ricerca del Senato Accademico dell’Università
di Verona (dal 1997 al 2001);
Tutorato agli studenti interni per
lo svolgimento del lavoro sperimentale ed anche correlatore di tesi
di laurea di Biochimica Vegetale (nel 1998), di Biopolimeri (nel
1999) e di Biochimica (2002), controrelatore di tesi di laurea del
corso di laurea in Biotecnologie Agro-Industriali;
Componente del Senato Accademico Allargato dell'Ateneo di
Verona in qualità di rappresentante dei ricercatori della
Facoltà di Scienze MM.FF.NN:(dal 1997 al 2001 ed a partire
dal Novembre 2004)
Componente della Commissione
Permanente per la Didattica del Senato Accademico dell’Università
di Verona
Referente della Facoltà di
Scienze MM.FF.NN. in seno alla Commissione Disabili di Ateneo
Componente della Commissione
Paritetica
Didattica:
·
1999/2000 e 2000/2001 - Corso di Laurea in Biotecnologie
Agro-Industriali
·
Docente del Corso di Fisiologia Vegetale;
·
Docente del Corso di Biochimica Vegetale.
·
2001/2002, 2002/2003, 2003/2004 - Corso di Laurea in
Biotecnologie Agro-Industriali
·
Docente del Corso di Fisiologia e Biochimica Vegetali (A.A.
2001/2002, 2002/2003, 2003/2004), 8 CFU + 1 CFU ;
·
Docente del corso di Fisiologia Vegetale (vecchio ordinamento,
A.A. 2001/2002).
·
2004/2005, 2005/2006, 2006/2007, 2007/2008 - Corso di Laurea
in Biotecnologie Agro-Industriali;
·
Docente del Corso di Fisiologia e Biochimica Vegetali, modulo
di Fisiologia Vegetale, 4 CFU + 1CFU.
·
2006/2007, 2007/2008, 2008/2009 – Corso di Laurea specialistica
in Biotecnologie Molecolari e Industriali
·
Docente del Corso di Biosintesi e Maturazione delle Proteine, 2
CFU.
·
2007/2008, 2008/2009 - Corso di Laurea in Biotecnologie
Agro-Industriali;
·
Docente del corso di Chimica e Biochimica Analitica –
modulo di Biochimica Analitica, 2 CFU + 1CFU.
·
2008/2009 - Corso di Laurea in Biotecnologie Agro-Industriali;
·
Docente del corso di Biologia Cellulare, 4 CFU + 1CFU.
·
Docente del corso di Biologia Vegetale, 2 CFU + 1 CFU
·
dal 2009 ad oggi - Corso di Laurea in Biotecnologie
·
Docente e coordinatore del corso di Biologia Generale e
Cellulare, 12 CFU.
ATTIVITA’ SCIENTIFICA - Anni Accademici
dal 1997 ad oggi
Meccanismi
molecolari di fotoprotezione
Il mio lavoro
di ricerca nel triennio 1997-2000 ha avuto come tema principale lo
studio del ruolo strutturale e funzionale dei carotenoidi nelle
proteine antenna delle piante superiori. L’importanza di queste
molecole è legata al ruolo chiave che giocano nei processi di
fotoprotezione nelle piante superiori. Il meccanismo principale sotto
indagine ha riguardato il non-photochemical quenching (NPQ)
dell’energia di eccitazione. Questo meccanismo è
dipendente dalla de-epossidazione della xantofilla violaxantina a
zeaxantina. La localizzazione di questi pigmenti nelle subunità
dei complessi del fotosistema I e II è di fondamentale
importanza nello studio del meccanismo molecolare della
fotoprotezione. Le ricerche svolte negli anni precedenti hanno
evidenziato la localizzazione della violaxantina (legata) nelle tre
subunità minori del fotosistema II chiamate CP29, CP26 e CP24
e di violaxantina associata debolmente nell’antenna maggiore
LHCII. La zeaxantina, d’altro canto, è stata localizzata
quasi esclusivamente legata alle antenne minori. Con una metodica,
messa a punto in questo laboratorio, è stato possibile
preparare proteine ricombinanti che legano tutti i pigmenti
normalmente presenti nelle proteine antenna native, ovvero
ricostituire in vitro le apoproteine sovraespresse in E.coli
con estratti di pigmenti di membrane fotosintetiche. Il risultato più
interessante è che i campioni ottenuti posseggono tutte le
caratteristiche funzionali (studiate mediante emissione di
fluorescenza in stato stazionario e risolte nel tempo, spettri di
assorbimento, ecc.) e strutturali (dicroismo circolare, dicroismo
lineare, stabilità, ecc..) riscontrate nei sistemi nativi.
Queste qualità e la versatilità del metodo di
ricostituzione in vitro hanno aperto la strada allo studio della
funzione e della localizzazione all’interno dei sistemi antenna
dei singoli carotenoidi. I risultati ottenuti hanno fornito
importanti delucidazioni sul diverso ruolo dei carotenoidi
all’interno delle antenne minori CP26 e CP29 (Figura1).
Dal punto di vista strutturale la luteina e la violaxantina sono
risultate le specie capaci di stabilizzare maggiormente la struttura
proteica. La ricostituzione con zeaxantina ha anch’essa dato
risultati positivi fornendo così un prezioso sistema per la
comprensione del ruolo funzionale di questa xantofilla che è
noto essere coinvolta direttamente nel processo di fotoprotezione
(ciclo delle xantofille). Quando di utilizza la sola neoxantina,
d’altro canto, non è possibile ottenere un sistema
ricostituito stabile; questo risultato dipende dalla stringenza della
metodica utilizzata che permette di selezionare la popolazione più
stabile di proteina ricostituita con pigmenti. Le misure dei tempi di
vita di fluorescenza delle clorofille legate da questi sistemi
antenna e la resa quantica di fluorescenza della clorofilla a,
in collaborazione con il Prof. Alfred. R. Holzwarth (MPI-Muelheim,
Germania) e con il Prof. Harry Frank (Storrs, U.S.A.), hanno
evidenziato il diverso ruolo funzionale delle xantofille legate da
queste proteine nei processi di raccolta della luce e di dissipazione
dell’energia di eccitazione.
Modello
strutturale di CP29 ed orientamento dei dipoli delle clorofille
L’omologia di sequenza tra le regioni transmembrana delle
antenne minori ed LHCII (di cui è disponibile la struttura
risolta a 3.4 Å con cristallografia elettronica) ci ha permesso
di costruire un modello strutturale di CP29 e di verificare la
posizione dei residui coinvolti nella coordinazione delle clorofille
(Figura2).
A partire da questo modello 3D ed utilizzando mutanti sito-specifici
su questi residui (dai quali si sono ottenuti gli spettri di
assorbimento e gli spettri di dicroismo lineare), è stato
possibile calcolare l’orientamento dei momenti di transizione
dipolare delle singole clorofille legate (Figura
3)
Ca2+-binding
Si è studiato il legame del Ca2+ con la
proteina antenna CP29. La marcatura con 45Ca2+
ha dimostrato che questa proteina è in grado di legare calcio.
Studi spettroscopici della proteina purificata, dopo aver sostituito
lo ione calcio con Itterbio (Yb3+) hanno confermato la
presenza di un sito specifico di legame. La posizione di questo sito
è stata identificata analizzando la sequenza primaria ed il
modello strutturale tridimensionale: il sito è localizzato nel
loop tra la prima e la seconda elica transmembrana, in una regione
esposta al lume tilacoidale. Il legame del calcio potrebbe essere
reversibile in funzione del pH rendendo la proteina un sensore dello
stato energetico sulla membrana: Il distacco del calcio, quindi,
funzionerebbe da switch per attivare i meccanismi di dissipazione
dell'energia.
PsbS
Recentemente, si è trovata un'altra proteina che sembra
essere direttamente coinvolta nei meccanismi di dissipazione
dell'energia (non photochemical quenching - NPQ) nel PSII: la
subunità PSII-S (o PSbS). I mutanti di Arabidopsis che mancano
del gene nucleace che codifica per PsbS è incapace di
rispondere rapidamente all'eccesso di luce nonostante il ciclo delle
xantofille sia inalterato. Per chiarire il ruolo di questa proteina
nella fotoprotezione, abbiamo studiato le caratteristiche strutturali
e funzionali e di legame con i pigmenti fotosintetici della proteina
nativa e della proteina ricombinante espressa in E. coli. I risultati
ottenuti mostrano che questa proteina non è in grado di legare
stabilmente i pigmenti fotosintetici. I meccanismi molecolari di
dissipazione devono quindi essere legati ad altre caratteristiche di
questo sistema proteico. L'ipotesi da noi avanzata prevede un
cambiamento conformazionale di questa proteina indotto
dall'acidificazione del lume; questo cambiamento influenzerebbe il
trasferimento dell'energia all'interno del PSII verso il centro di
reazione e causerebbe un accumulo dell'energia di attivazione sulle
antenne minori collocate in prossimità di PsbS. I risultati
della marcatura di PsbS con 14C- DCCD sono a favore di
questa ipotesi funzionale.
Plant
Lipid Binding Proteins (nsLTPs) e apoptosi
Le
LTP (lipid transfer proteins) sono piccole proteine basiche presenti
in molti tessuti vegetali; sono state identificate
originariamente per la loro capacità di catalizzare lo scambio
di lipidi tra membrane in vitro. In seguito, il ruolo proposto di
scambiatori di lipidi intracellulari è stato messo in
discussione poichè sono state trovate anche nell'ambiente
extracellulare. Recentemente è stato proposto anche che
possano avere un importante ruolo nei meccanismi di difesa da
patogeni delle piante.
Queste proteine sono molto studiate anche
per la loro capacità di indurre forti reazioni allergiche.
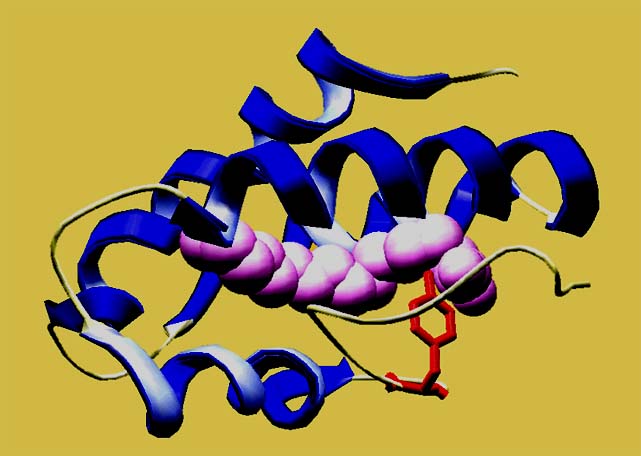
La
struttura tridimensionale di queste
proteine è nota: tutte sono formate da quattro alfa-eliche
tenute assieme da ponti disolfuro. Queste quattro eliche formano una
cavità idrofobica centrale nella quale può legarsi un
lipide. Le molecole lipidiche trovate legate sono molte e di diversa
natura, da acidi grassi a fosfolipidi. Il ruolo di questo legame e la
funzione svolta da queste proteine resta ancora motivo di studio
Il
progetto si propone di studiare le interazioni delle LTP estratte da
diversi sorgenti (semi di mais, semi di frumento) con le membrane
biologiche. Questo viene fatto utilizzando le LTP purificate, con
lipidi legati e senza.
Si utilizzano membrane artificiali
(liposomi) e membrane naturali (mitocondri) per verificare sia
l'azione aspecifica sul doppio strato lipidico sia l'azione
specifica, quella mediata da sistemi di membrana. Questo ultimo
aspetto è di grande interesse perchè può fornire
importanti informazioni sui meccanismi che controllano l'instaurarsi
della risposta apoptotica dei mitocondri.
Si è infatti rovata una regione omologia di sequenza tra le
LTP e la proteina pro-apoptotica Bid. Questa similitudine non si
limita alla sequenza: è stato visto sperimentalemnte che Bid
può legare e scambiare molecole lipidiche. Queste molecole
potrebbero quindi agire come segnale per l'attivazione
dell'apoptosi.
Su questo argomento di ricerca è stato
presentato un poster al Congresso  Clicca il logo per visualizzarlo.
Clicca il logo per visualizzarlo.
Bid
e la scelta di attivare la morte programmata - Apoptosi
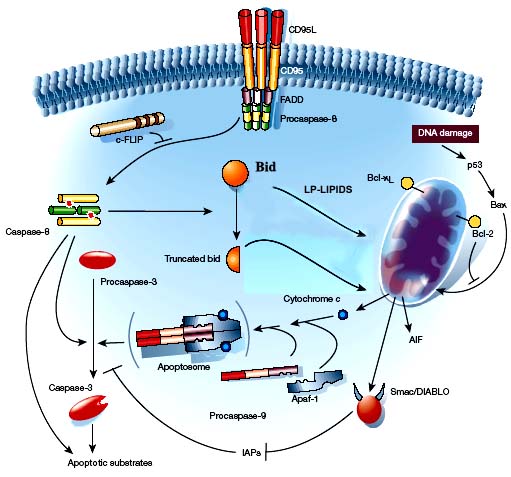
La proteina pro-apoptotica Bid si trova al crocevia delle due vie
di attivazione cel processo apoptotico: la via intrinseca e la via
estrinseca. Questa proteina è estremamente importante poiché
agisce da sensore e può scatenare a valle l'attivazione delle
proteine effettrici dell'apoptosi, quelle che agiscono direttamente
sulla membrana esterna mitocondriale causando i cambiamenti
irreversibili che scatenano l'esecuzione del processo di distruzione
controllata della cellula. Lo studio delle caratteristiche
strutturali e funzionali di questa proteina è di cruciale
importanza per comprendere come si possa influenzare la scelta di
attivare la morte programmata in cellule che hanno perso il controllo
normale. Caratterizzare infatti i ligandi specifici (molecole
lipidiche e derivati, quali i lisolipidi) e il sito di legame nella
proteina ci potrà permettere di sviluppare nuove molecole in
grado di influenzare il processo apoptotico. La proteina Bid
ricombinante è anche utilizzata per studi di interazione
proteina-proteina con altri membri della famiglia Bcl-2 per studiare
il meccanismo di interazione con le proteine anti-apoptotiche
attraverso anche il tentativo di cristallizzare il complesso per
determinarne la struttura tridimensionale.
back
to the home page
{kind=link}
{kind=link}
{kind=link}
{kind=link}